Table of Contents (click to expand)
Worker bees aren’t truly sterile. The queen’s pheromones switch off their ovaries, so they raise her young instead of laying their own. This pays off genetically: because bees are haplodiploid, a worker can be more related to her sisters than to her own offspring. Remove the queen and some workers do lay eggs, but only unfertilized ones that become male drones.
In 1831, Charles Darwin set sail on the HMS Beagle for a five-year voyage around the world, having abandoned a half-hearted run at a medical career to chase his real passion: natural history. On that trip, which took in the Galapagos Islands, he had one of the most important intellectual epiphanies in human history – he began conceptualizing his theory of natural selection. Another man, Alfred Russel Wallace, stumbled upon a similar epiphany in the Amazon rainforest while collecting exotic bird species. These two men laid the first foundations of evolution and natural selection as we know them today.
However, there was one tiny glitch in this nearly perfect tale. Darwin couldn’t figure out why, in some animals such as bees, only one female reproduces while the rest tend her brood instead of raising young of their own. If natural selection rewards passing on your own genes, why would workers give that up? Darwin never fully cracked this puzzle in his lifetime, and it took many more years and numerous valiant attempts to reach a satisfying answer. To get there, we first need to understand what biologists mean by reproductive fitness.
Reproductive fitness and mating
In the natural world, reproduction is all important. Living beings wage war just to woo the opposite sex, as the ultimate goal is to get as much of one’s genome into the next generation as possible. To achieve this, animals work to demonstrate that their DNA is superior and should be perpetuated.
Darwinian Fitness
Since plants and (most) animals lack microscopes or the luxury of performing DNA sequencing, other observable strategies have been employed. There are flamboyant colors, elaborate dances, big muscles, perfumery and loud, deafening roars. All of this is done in the great pursuit of spreading as many of their genes as possible on to the next generation. This is what evolutionary biologists call Darwinian fitness, relative fitness or reproductive fitness (these terms are often used interchangeably).
Relative fitness is formally defined as the relative probability that a hereditary characteristic will be reproduced. In simpler terms, it is the likelihood that a certain characteristic will be passed down; genetically speaking, it is the probability that an allele of a gene will be expressed in future generations. At the most fundamental level, natural selection can be thought of as the process by which genes can successfully survive and thrive.
Inclusive Fitness Theory
Distributing the greatest number of your genes into the population can be done in one of two ways, individually, by having many offspring, or altruistically, by increasing the chances that a family member sharing part of your DNA will have many offspring. In other words, you can either increase your own fitness or increase your kin’s fitness. This is called inclusive fitness theory.
To explain this further, we’ll use an analogy from J.B.S. Haldane, the renowned geneticist and biochemist. The story goes that Haldane was once sitting with a graduate student of his in a bar when he was struck by a thought. Haldane performed some quick calculations and stated that, if only one or two of his brothers were drowning in a river, he would not bother to save them, but if more than two were drowning, he would risk his own life to rescue them. This story may or may not be true, and various versions of the tale exist, but it illustrates a key concept.
You share 100% (1.0) of your DNA with yourself. When you reproduce, each of your offspring will get 50% (0.5) of that DNA. The more offspring you have, the higher the percentage of your DNA entering the gene pool. With your siblings, you share 50% (0.5) of your DNA and 25% (0.25) of it with any nephew or niece. Therefore, caring for one sibling or 2 nieces is ostensibly the same as caring for one of your own children!
Therefore, saving 1 sibling (0.5×1=0.5) or 3 nephews/nieces (0.25×3=0.75) is less than 1.0, so you will lose part of your genome if you save them at the expense of your own life. However, if 3 siblings are drowning, it would be worth saving them, as they will have a higher chance of propagating your DNA to their offspring (0.5×3=1.50). Similar calculations would reach the same conclusion for 5 nieces/nephews (go ahead, work it out for yourself).
| Selfishness | Altruism |
| You at the bank of the river = 100% or 1 1 brother drowning = 50% or 1/2 Do not attempt to save | You at the bank of the river = 100% or 1 3 brother drowning = 150% or 3/2 Attempt to save |
| You at the bank of the river = 100% or 1 3 nephews drowning = 75% or 3/4 Do not attempt to save | You at the bank of the river = 100% or 1 5 nephews drowning = 125% or 5/4 Attempt to save |
Tabular representation of Haldane’s analogy.
Haldane said something to this effect in an article published in the now defunct magazine Penguin New Biology in 1953.
Although this illuminated part of the mystery surrounding the peculiar reproduction of social insects, it did not provide a complete and utterly satisfactory scientific answer for why female worker bees were sterile. At least, it wasn’t clarified until W.D. Hamilton entered the scene.
W.D. Hamilton was a British evolutionary biologist and geneticist who made one of the most important advancements in evolutionary biology. In 1964, Hamilton published a wordy and relatively mathematical paper (at least for evolutionary biology at the time) titled “The Genetical Evolution of Social Behaviour”. This paper, despite its lack of eloquence, solved Darwin’s conundrum both mathematically and elegantly.
The Solution: Hamilton’s Rule

Hamilton’s Rule.
The above picture shows Hamilton’s rule and the mathematical formula in a simplified form.
Hamilton’s rule simply states that a particular social behavior will be favored if b, the benefit to the recipient (in terms of reproductive fitness), is greater than c, the cost born by the donor (in terms of reproductive fitness). Mathematically, this can be expressed as rb > c, where r is the coefficient of relatedness that measures the statistical relationship between the donor’s genes and recipient’s genes.
How does Hamilton’s rule tell us anything about social insects and sterile worker bees?
Before explaining how Hamilton’s rule explains worker bee sterility, we must take a quick look into bee reproduction and the distribution of genetic material amongst the male and female gametes.
In humans, there is a simple halving of genetic material, 23 chromosomes from the female and 23 chromosomes from the male. This isn’t the case in bees, as they are haplodiploid creatures, meaning that the female has 2n chromosomes, but the male has only n (half) chromosomes. This changes the relatedness that any offspring will have to each other and to their parents.
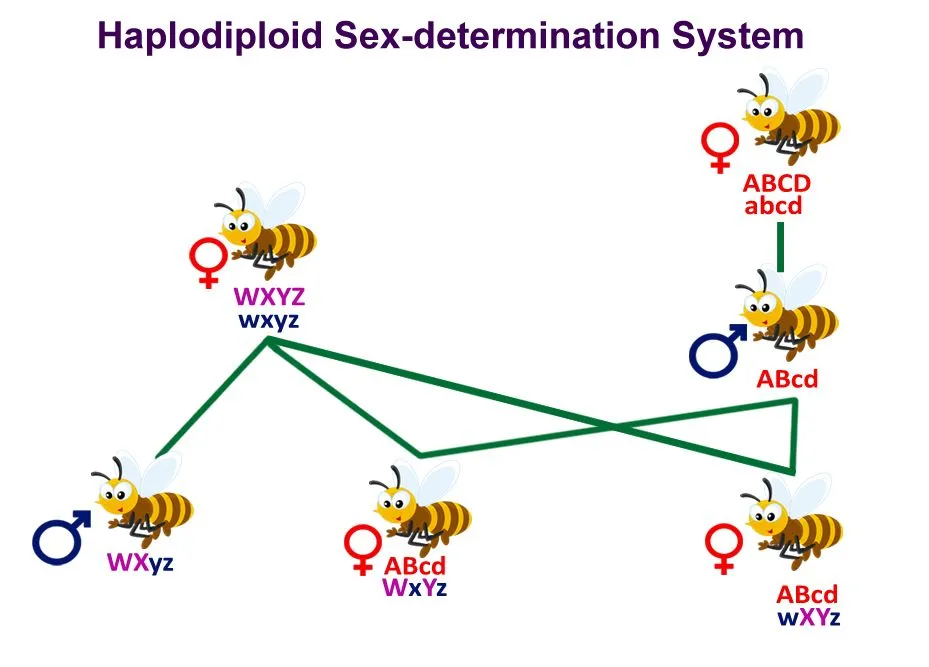
Haplodiploidy in bees.
Daughters are related to their mother by r = ½, but because a father is haploid and passes his entire single set of genes to every daughter, full sisters are related through their father by r = 1. Averaging the two parents (and assuming the queen mated with a single male) gives sisters a relatedness of r = ½(1) + ½(½) = ¾, higher than the r = ½ they would share with their own offspring.
Since the female workers would only be related to their offspring by r = ½, the benefit of rearing their offspring and increasing the reproductive fitness of the queen is greater than the cost. A female worker is actually helping establish her own genome in the gene pool better, as she shares more DNA with her sibling than she would with her offspring.
There is a catch, though. That tidy three-quarters figure only holds if the queen mates with a single drone. Real honey bee queens are famously promiscuous, mating with a dozen or more drones on their nuptial flights, so a hive is really a patchwork of half-sisters. Once you average across all that mixed paternity, the relatedness among workers in a typical colony falls to roughly 0.25–0.3, below the 0.5 they would share with their own sons. So the elegant 3⁄4 story cannot be the whole explanation.
Hamilton’s rule itself remains a cornerstone of evolutionary biology, but the specific “haplodiploidy hypothesis” (that the 3⁄4 sister relatedness is what drove worker sterility) has not held up well. When biologist Jacobus Boomsma and others looked closely, two problems stood out. First, because queens mate with many drones, real relatedness is far below 3⁄4. Second, sterile workers have also evolved in plenty of animals that are not haplodiploid, such as termites, where both sexes are diploid. A 2014 comparative analysis of thousands of insect species found no sign that haplodiploidy by itself speeds up the evolution of sterile castes.
The factor that now looks decisive is lifetime monogamy. If a mother mates with a single male for life, a helper is exactly as related to her siblings as she would be to her own offspring (both 0.5), so the relatedness terms cancel out of Hamilton’s rule and even a tiny efficiency gain from helping is enough to favor sterile workers. So the modern answer keeps Hamilton’s elegant math but trades the haplodiploidy story for a monogamous ancestry, with plenty of molecular and ecological details still being worked out.
So Are Worker Bees Actually Sterile?
All of the above explains the evolutionary why. The day-to-day how is a separate, and rather neater, story, and it is the reason calling workers “sterile” is a bit of a fib. A worker bee is born with perfectly good ovaries. They are simply switched off.
The off switch is chemical. The queen constantly releases a blend of compounds called queen mandibular pheromone (QMP), which her attendants spread through the hive as they groom and feed one another. Together with a pheromone given off by the developing brood, QMP suppresses the workers’ ovaries so that almost none of them ever lay an egg. Even so, the suppression is not perfect: in a healthy, queenright colony, roughly 1% of workers have ovaries developed enough to produce eggs.
Take the queen away, and the spell breaks. Once a colony has been queenless (and broodless) for around three weeks, the pheromone signal fades and a handful of workers become laying workers. Here is the twist: a worker has never mated, so she can only lay unfertilized eggs. Thanks to haplodiploidy, those eggs can only become haploid males, so a laying worker produces nothing but undersized drones. She literally cannot make a daughter, let alone a new queen, which is why a queenless hive that reaches this stage is usually doomed.
To keep cheats in check while the queen is alive, other workers act as a police force, hunting down and eating any worker-laid eggs they find, a behavior aptly named worker policing. So the honest answer to the headline question is that female worker bees are not strictly sterile at all. They are reproductively suppressed, by chemistry while the queen reigns and by their own sisters if they try to break the rules.
References (click to expand)
- Boomsma, J. J. (2009). Lifetime monogamy and the evolution of eusociality. Philosophical Transactions of the Royal Society B. PubMed Central (PMC).
- Hamilton, W. D. (1964, July). The genetical evolution of social behaviour. II. Journal of Theoretical Biology. Elsevier BV.
- Gadagkar R. (2001). Survival Strategies: Cooperation and Conflict in Animal Societies. Harvard University Press
- The Colony and Its Organization. Mid-Atlantic Apiculture Research and Extension Consortium (MAAREC), University of Delaware.
- Reproductive parasitism by worker honey bees suppressed by queens through regulation of worker mandibular secretions. Scientific Reports. PubMed Central (PMC).